


研究内容
本研究室における研究の原動力は、野外で観察される多様な種が進化してきた謎への好奇心です。基礎科学研究を行うそもそもの動機は、身の回りの現象への素朴な疑問と好奇心であると思います。それを一連の確立された手順で解き明かしていくのが科学であると考えます。
私の専門とする生態遺伝学とは、1964年にEcological Geneticsの本を著したE.B.Fordの序文によると「野外調査と実験室内での遺伝学を融合した手法を用いて行われる進化と適応に関する実験的研究」です。すなわち、野外で進行する生物進化の遺伝機構を研究する学問といえます。現在、もっとも興味をもって研究しているテーマは「種分化を完成に導く要因」と「新しいニッチへの進出のメカニズム」です。種分化とは、集団間に生じる遺伝的な違いによって、互いの遺伝子流動(遺伝子の交換)が低下する過程です。すなわち、連続的な変異から非連続的な単位としての種が生まれる過程であり、種分化は生命が示す本質的な性質の一つといえます。私の研究室では、生態適応などで開始した種分化がどのように完成に至るのか、その過程を研究しています。また、生物は地球上の様々な環境に進出し適応してきました。この適応進化も生命が示す本質的な性質の一つです。しかし、新たな環境に適応して分布を広げることができる種とできない種がいます。この違いは一体どのような要因によるのでしょうか?イトヨは様々な淡水域に進出して適応放散を遂げましたが、近縁の二ホンイトヨは海にとどまっており淡水魚にはなれていません。この違いの原因を探っています。
そのために我々は、表現型多様化や種分化の原因となる遺伝子、さらには原因変異の特定を目指しています。原因遺伝子、さらには原因突然変異が特定できれば、原因突然変異の数や効果の推定、原因突然変異の起こりやすさなどを理解することが可能となり、野外生物の進化をめぐる上記の問いの解明につながると考えるからです。これらを一つ一つ地道に解き明かしながら、いつか種分化の統一的理論のようなものに辿り着ければと思い、日々研究しています。
このゴールを達成するために、下記のような一連の研究手法を確立してきました。
(1)まず、野外より魚を採集し、集団や種間での形態や生理機能の違い、雑種で生じる異常などを出来る限り詳細に観察し記載します。地道な観察こそが重要な第一歩と考えています。
(2)ついで、その生物の適応度にとって重要と考えられる形質の候補原因遺伝子について、QTL解析、全ゲノム比較、トランスクリプトーム解析、クロマチン解析などを用いて探索します。
(3)得られた候補遺伝子について、生化学的/分子生物学的な解析を試験管内で行ったり、ノックアウトやトランスジェニック魚を利用して生体での機能解析を行ったりすることで、原因遺伝子の特定を行います。
(4)さらに、原因遺伝子座の野外での振る舞いについて、理論集団遺伝学的解析やコンピューターシミュレーションで解析したり、遺伝研所内の人工池を用いたりして明らかにしていきます。
これら一連の手法は、拙著「生態遺伝学入門(丸善出版)」にまとめさせて頂きました。苦手な手法については、国内外のグループと必要に応じて柔軟に共同研究を行ったり、新しい手法や考え方を学んでいきます。
これら作業の連続はとても時間のかかる地道なものです。しかし、国内外の理解しあえる仲間たちと知恵を出し合い、協調性をもって一つ一つ未知の扉を開いていく過程を、わくわくドキドキしながら一喜一憂しながらできる人には、私の研究室は向いているかもしれません。この試行錯誤の過程で見つけた発見は、どんなに小さな発見であっても嬉しくて、それを発表して世界が驚いてくれて、世界中で似た研究が続いていけば、その喜びはなおさらです。モットーは、「本当に面白いね!」と言ってもらえる研究、世界が真似をするような研究を自分のラボから世界へ発信することです。世界初のことをやったら、きっと自然界が何か新しいメッセージを見せてくれるはずと信じて研究しています。自分の論文が(もちろん現在もですが)数十年後、あるいは、百年後にでも世界のどこかで誰かに読まれて、刺激を与え続けることができたらいいなと思います。
自分なりにとても気に入っている論文はどれも当初の予定通りではなく、予想外の展開が複数あったものです。今、論文を読み返してみても、自分自身や共同研究者との一喜一憂が思い出されます。予想通りの結果というのは、自分の頭の中で思いつくこと、あるいはAIが思いつくことにすぎません。予想外の結果というのは自然界が我々に何かを語りかけている瞬間で、実は以外に多いはずなのですが、出会ってもその重要性に気づかなかったり、忙しいことを言い訳に目をつぶってしまうことが圧倒的に多いと思います。なにかの発見をするためには、自分とは異なる考え方を受け入れ、いつもオープンな気持ちで予想外との出会いを待ち受けている姿勢が必要だと常々感じています。
進行中の研究課題
トゲウオ科のイトヨは、約200万年以内に多様化を遂げたことから、生物多様性進化の研究の格好のモデル生物です。また、メダカ科魚類は、日本を含む東南アジアにおいて著しい多様化を遂げています。これらイトヨやメダカを利用して以下の課題に迫ります。特定の魚で見出した原理の普遍性については、メタ解析で検証します。
1.種分化の遺伝機構
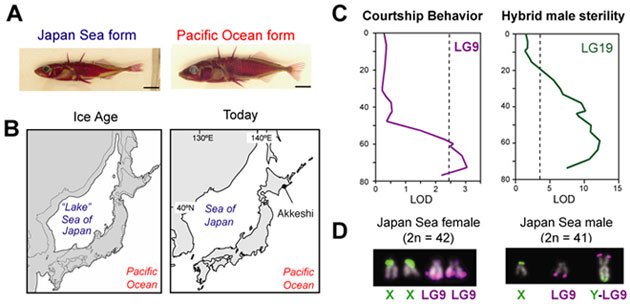
種分化、すなわち、近縁種間での生殖隔離機構の機構に迫ります。日本には、日本海型と太平洋型の二種のイトヨが生息しています。これら二種は性染色体転座 など核型の点で異なるのみならず、求愛行動も異なっておりランダムには交配できません。また、無理にかけ合わせても交配方向によっては、雑種オスが不妊になります。連鎖解析やゲノム解析の結果、ネオ性染色体の進化と種分化に深いつながりがあることが分りました。これら二種のイトヨの生殖隔離の原因遺伝子に迫 ります。
2.新規ニッチへの進出の遺伝機構
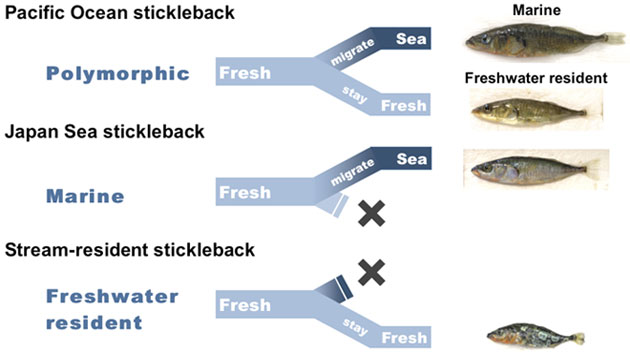
新しいニッチへの侵入は、その後の適応放散を誘導する場合があります。トゲウオは、淡水域に進出することで適応放散を遂げました。しかし、新規ニッチへ侵入できた分類群もあれば、できなかった分類群も存在します。これら、新規ニッチへの適応進化の違いを生み出す原因となる遺伝基盤について、異なる塩分適応能や異なる代謝能力をもつイトヨ3型を用いて研究します。また、ホルモン内分泌系の進化は、複数形質の同時的な変化を引き起こしうることから、新規環境への適応に重要ではないかという仮説のもと、エコタイプ間のホルモン分化の機構も明らかにします。
3.性染色体ターンオーバーが進化に果たす役割
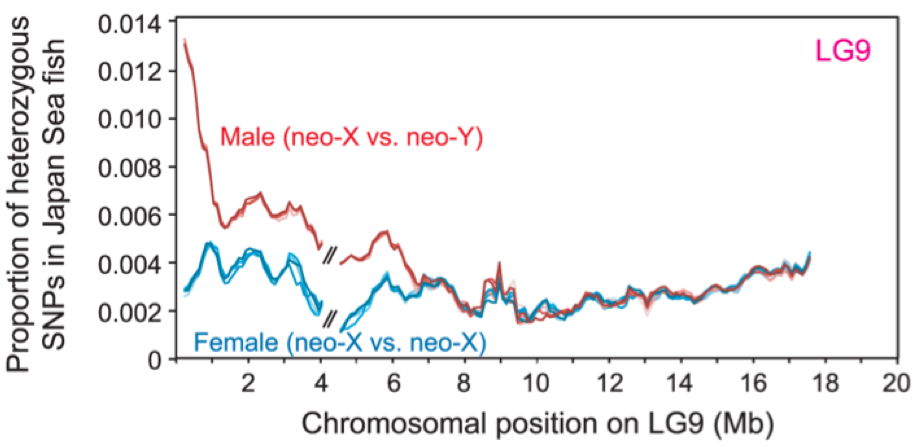
性染色体は、ゲノムの中でもとりわけ進化速度の速い領域で、魚類などでは近縁種間でも性染色体が異なっていることは多々あります。我々は、こういった性染色体のターンオーバー(転換)が、種分化や性的二型の多様化に果たす役割について、ゲノム解析、遺伝的連鎖解析、候補遺伝子の同定、集団遺伝理論解析などを 通して解明します。
4.進化生物学の応用的側面
人為的環境改変は急速に環境を改変しており、生物に与える影響が懸念されています。また、津波をはじめとする環境変動も野外生物に影響を与えると考えられます。さらに、外来種は新規環境へ適応しながら拡散し、社会問題となっています。急速な環境変化に対して生物がどう反応するのか、どのように絶滅リスクを軽減できるのか、また、外来種は如何にしてうまく新規環境に適応していくのかについて、イトヨをモデルにして研究しています。
